Rust disease of poplar and willow
Ian McIvor and Siva Sivakumaran, New Zealand Tree Grower November 2009.



Rust fungi found on poplars and also on willows belong to the genus Melampsora. Rust caused by Melampsora is one of the most important leaf diseases of poplars. Internationally some 13 species and two hybrids of Melampsora on poplars have been described, of which three have been identified in New Zealand, namely, Melampsora larici-populina, M. medusae and M. medusae-populina, a unique interspecific hybrid.
A challenge
Rusts pose the greatest disease challenge to willows and poplars in New Zealand. Rust fungi have complex life cycles involving up to five different spore stages. These fungi often need two unrelated host plants to complete their full life cycles. However they do not require a secondary host and can re-infect without going through all five stages.
The urediniospore stage is what is seen in the nursery. Urediniospores are capable of producing many cycles of the same form during the season, causing disease epidemics on poplars. Where leaf rusts are common and severe, poplar and willow leaves may become distorted, wither and drop prematurely reducing growth up to 30 per cent. The more susceptible a tree is to leaf rust the more rapidly the disease develops and the sooner defoliation starts.
Growth loss due to rust is often masked by the normally rapid growth of poplars and willows and can best be measured by comparing the growth of infected trees with comparable healthy trees protected by fungicide sprays. Continued defoliation of successive flushes of growth by leaf rusts decreases vigour and indirectly increases the likelihood of winter injury as well as susceptibility to other diseases, such as silverleaf fungus.
Spray early
Rust infection should be anticipated by starting a spray control programme two to three weeks before the infection normally occurs. The top photograph shows the location of the rust infection inside the leaf and the visible rust is due to the spores erupting through the leaf surface.
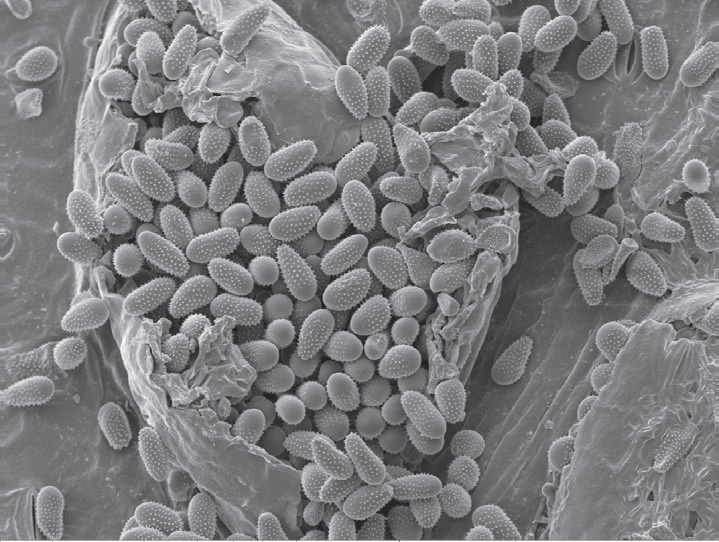

The surface of urediniospores is covered with spines. Rust species are identified from their location on the leaf either underside or both sides, and the size and spininess of their urediniospores. Urediniospores of M. larici-populina have a smooth surface at the apex, while M. medusae have a smooth surface on the equatorial zone. The hybrid has smooth areas at the apex and equator.
Assessment
A national assessment of the status of rust species has been set up to determine
- If any new species have arrived in New Zealand
- Whether further hybridisation has occurred between known rust species
- The relative frequency of the different species.
At this stage we have used scanning electron microscopy to study urediniospore structure for the different samples and identify the species from already published data. Poplar rust leaf samples were collected for different poplar clones and from nurseries in different regions, and the external spore structure analysed using scanning electron microscopy.
Urediniospores from all 17 samples evaluated had smooth apical patches as seen in the photographs.The length of the spores was around 25 μm, and width 12 μm.
| Clone Region | Veronese | Toa | Kawa | Fraser | P. tricho x P. nigra | Crows nest |
|---|---|---|---|---|---|---|
| Taranaki | x | x | x | |||
| Wairarapa | x | x | x | |||
| Hawke's Bay | x | |||||
| Wairoa | x | |||||
| Gisborne | x | x | ||||
| Manawatu | x | x | x | x |
All rust samples were identified as belonging to M. larici-populina. Urediniospores of M. medusae were identified in a previous survey carried out by the same researchers, but the present results suggest that M. larici-populina is most prevalent in the nurseries. More importantly there was no evidence of hybridisation or new rust species based on the urediniospore appearance. If secondary host trees are not close, the rust sexual cycle will not be completed, there will be no hybridisation and new rust infections will be from asexually produced spores which have a very low level of variability from generation to generation.
Resistance
It is easy to notice rust susceptible poplar clones in autumn. They have lost almost all their leaves except for a tuft of leaves at the very top not yet infected with the rust fungus. Many of the older poplar trees are susceptible, dating back to before the rust M. larici-populina arrived in New Zealand in 1973.
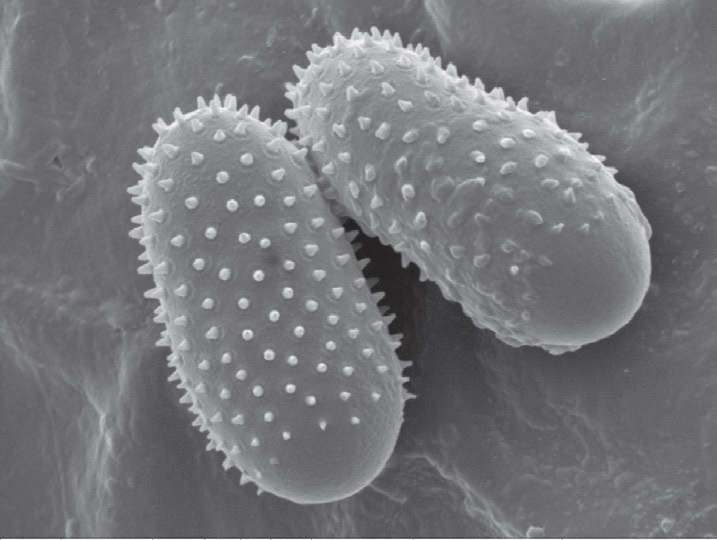
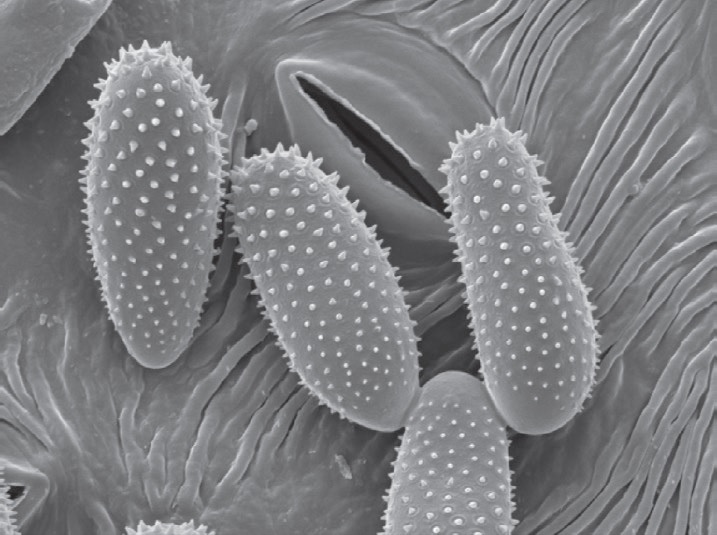
hole, the stoma
Poplar and willow clones bred and released after 1980 were selected for high resistance to rust and these show up on the landscape as still in green leaf in late autumn. Rust races evolve and it is likely that poplar and willow clones will become less resistant to rust. For this reason, new hybrids are being bred which are selected for resistance to the current strains of rust which may or may not be the same as previous strains. The time between initial selection and eventual commercial release is of the order 12 to 15 years. Fortunately research suggests rust races do not change rapidly.
When replacing old poplar and willow trees select those clones suitable for your region. Wetter climates favour rust, whereas drier climates inhibit rust spore germination. In addition to other characteristics such as vigour, form, fodder and timber potential it is important to replant with clones having a high rust resistance, particularly in wetter climates.
Ian McIvor and Siva Sivakumaran work for Plant & Food Research