Plantation silviculture at the crossroads
Euan Mason, New Zealand Tree Grower May 2007.
What do you think of New Zealand? The classic question reputedly asked of so many visitors has provoked a variety of answers. John Lennon replied that after 10 minutes on the ground he thought the inside of the terminal was just fine. I returned to New Zealand with a freshly minted forestry degree in 1975 after a globe-trotting childhood. Dave Elliott, then principal forester at Kaingaroa, waited a couple of weeks before posing the forestry equivalent question – What do you think of forestry in New Zealand?
I did not hesitate with my reply – You are not fully using the site.
Unique silviculture
There followed a careful explanation of what, at the time, made New Zealand plantation silviculture unique. New Zealand silviculturalists trade volume production for value in a highly productive environment, leading them to unusual silvicultural strategies with very low stocking rates. I explained this to an audience in the southern United States last year. When I got to the part about pruned regimes on low quality sites with final crop stocking rates of 80 stems per acre, a voice in the back of the room exclaimed that was ridiculous. Many people in the United States still equate volume with value.
Until recently, value in New Zealand was defined by large scale log geometry, with small defect cores, large stem diameters, small branches on unpruned logs and minimal sweep. All this was paramount in the minds of decision makers as they crafted their silvicultural regimes.
Farm foresters leading the way
Some researchers point out that radiata pine was never intended to produce structural products, and that the features that make it an excellent finishing timber reduce its utility as a structural one. Others would prefer that we switched to alternatives such as eucalypts for producing structural wood. They may be right, and perhaps farm foresters who frequently experiment with alternative species, will lead the way.
New objectives for growing plantations cause us to redefine silvicultural strategies, and make silvicultural stand models obsolete.
Research at the School of Forestry aims to support forest growers as they decide how to manage their future crops in the face of changing objectives. If we also consider influences of climate change and carbon sequestration, then our silvicultural strategies will change more.
New definitions

We have witnessed a dramatic change in focus over the last few years. Late last year I took a group of forestry students on a trek into the wilds of the Maruia, to the mainland island at St Arnaud and then on to Nelson. Here they learned about forestry from staff at Rayonier, Weyerhaeuser and Nelson Pine International. Nelson Pine sets a velocity limit for logs, measured with a resonance acoustic device, of 3.1 km per second. Velocity is correlated with wood stiffness, an essential feature for structural lumber and wood composites. New definitions of value require changes in silvicultural thinking and new kinds of assessments. The initial plea from log buyers has been for longer rotations.
Older trees, it was reasoned, would contain lower proportions of core wood – wood growing close to the pith. Compared to outer wood, core wood has lower density, higher microfibril angles, higher gradients in the microfibril angles and consequently higher gradients in longitudinal shrinkage. These features make the wood less stiff, and less stable during drying, than outer wood. However, long rotations come at a cost, and so there have to be better ways to improve value.
Breeding the answer?
Breeders began to measure density and log velocity, thought to be a measure of average microfibril angles, noting that many wood properties are highly heritable. Breeding and perhaps clonal forestry can be part of the answer, but silviculture is at least as important as genotype.
Arturo Bascunan, a Chilean student at the New Zealand School of Forestry, showed that stems closer to stand edges had lower velocities, suggesting that sway may influence microfibril angles.
Another Chilean forestry student demonstrated that higher initial stockings of radiata pine promoted growth of core wood with higher acoustic velocities, and more recently Matt Waghorn, also a forestry student, mapped log velocity up stems of different breeds at a range of stockings. He noted that while the first two metres of a stem often had the lowest velocity, the section from two to four metres above ground level often had the highest velocity.
Presumably, if high velocity logs are sufficiently more valuable than lower velocity ones, managers might increase crop value by lopping off the lower sections of stems before buckling. Mike Watt of Ensis noted that stem slenderness was often related to velocity at breast height, and pointed to a functional relation that might compel a slender tree to become stiffer in order to prevent stem buckling. Matt’s results appear to show that the ‘avoidance of stem buckling’ hypothesis cannot explain patterns of log velocity observed up stems. Some of my personal research indicated that distance to canopy might also influence velocity, corroborating some theories of P R Larson from the 1960s who suggested auxins exuded by new buds and foliage might influence microfibril angles and therefore velocity.
No interaction
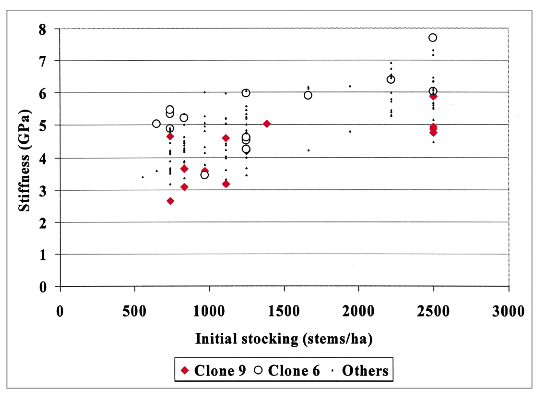
One fascinating theme running through all our work so far is that genetic and silvicultural effects on velocity do not interact – they are additive. Whatever genotypes are doing to alter log acoustic velocity, it is unlikely to comprise the same processes that drive silvicultural effects.
These theories do not explain why wood grown in Canterbury lacks stiffness. In addition, some of my research found that competition from grass in Canterbury resulted in lower stiffness, unlike between-tree competition which increased stiffness. For a possible explanation we need to turn to some research conducted in the Pacific Northwest by Domec and Gartner. They compared effects of core wood formation on structural stability with effects of core wood on plant water relations. They showed that some important features of core wood allow trees to minimise cavitation – the entry of air into their water transport systems high in a stem. Dry regions like Canterbury may grow less stiff wood because water stresses are high.
Why chase explanations
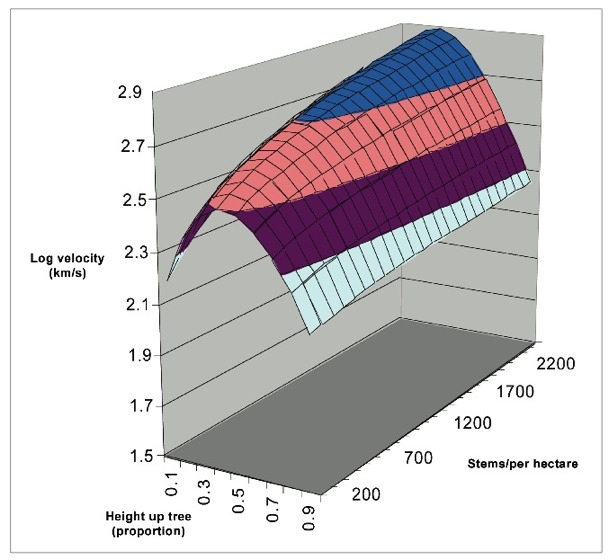
Why are we chasing all these explanations? Partly for the joy of discovery, but mostly because forest growers are asking two questions in the face of changing definitions of value for structural logs –
- Where and which are my high velocity stems?
- How do I create higher velocity ones in future?
They need models that represent where their stiff wood is and decision support systems that give silviculturalists feedback about the nature of the wood they are creating. Moreover, in my view we need models of wood properties at a ring level, because gradients in microfibril angles and density affect stability of sawn lumber.
Why not simply include measured effects of stocking and genotype directly in our models? The reason why this might be misleading is that many explanatory variables are collinear in stocking experiments, or to put it more simply, they vary together. For example as stocking increases then radial growth rate drops, canopies move up stems more rapidly over time, wind loading on each stem is less and stems become more slender.
Much to learn

Suppose we build a ring level model of microfibril angle development using radial growth rate as an independent variable, and a manager decides to reduce core wood microfibril angle by retaining weeds on a site, thereby restricting radial growth. It would make sense, and the model would indicate that log velocity should increase. However, in my experiment at Dunsandel, trees subjected to the most competition from grass had the lowest velocities. Clearly there is much we need to learn before we can produce robust silvicultural models that include log velocity outputs, let alone estimates of stability during drying.
When we produce such models, we will need a structural log index equivalent in function to Jim Park’s pruned log index that relates measurable log features to log value in a structural log market. Log velocity might give us an index of stiffness, but is it adequate as an index of stability during drying? We clearly need log and branch dimensions in the structural log index, and I suspect we might need to add something more elaborate. For example, the volume weighted mean gradient in microfibril angle within 50 mm segments of radius within a log, given that much structural lumber is cut to 50 mm by 100 mm.
Euan Mason is Associate Professor, New Zealand School of Forestry, University of Canterbury