

PESTS AND DISEASES OF FORESTRY IN NEW ZEALAND
Poplar leaf rusts caused by Melampsora spp.
Scion is the leading provider of forest-related knowledge in New Zealand
Formerly known as the Forest Research Institute, Scion has been a leader in research relating to forest health for over 50 years. The Rotorua-based Crown Research Institute continues to provide science that will protect all forests from damage caused by insect pests, pathogens and weeds. The information presented below arises from these research activities.
Poplar leaf rusts caused by Melampsora spp.
From Scion publication Forest Research Bulletin 220,
An Introduction to The Diseases of Forest and Amenity Trees in New Zealand,
G.S.Ridley and M.A. Dick 2001.
Species: Melampsora larici-populina and M. medusae (Basidiomycete)
Common name: Poplar leaf rust
Country of origin: Melampsora larici-populina is from Europe and M. medusae is from North America.
Host(s): Uredinia of Melampsora larici-populina occurs on many poplar species and clones; spermogonia and aecia are formed on Larix decidua and L. kaempferi, and very rarely on Pinus radiata.
Melampsora medusae forms uredinia on many poplar species and clones but not the semi-evergreens; it does not infect conifers (Fig. 52).
Symptoms: Small orange-yellow pustules on one or both leaf surfaces; in severe infection, trees have a golden appearance; heavily infected leaves turn brown, wither, and curl at the margin before falling.
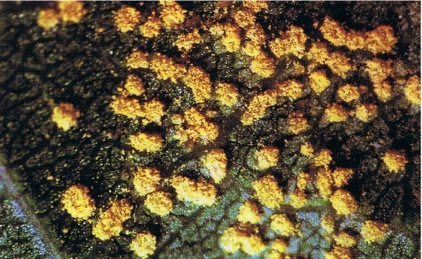
Fig. 52: Rust pustules of Melampsora medusae on the leaves of a Populus hybrid

Fig. 53: Rust pustules of Melampsora larici-populina on the underside of a Populus hybrid leaf
Disease development: The most visible parts of the cycle for poplar rusts caused by Melampsora larici-populina are the urediniospores (Fig. 53). The urediniospores are wind blown, infecting further poplars, and producing a new crop of urediniospores every 2 weeks. Free water on the leaf surface is needed for urediniospore germination and host infection. On semi-evergreen poplars the fungus can continue to produce urediniospores all through winter and spring and can therefore survive without producing any other spore stage.
On older infected leaves, teliospores are produced; these remain dormant on fallen leaves until the following spring when they germinate to produce basidiospores which can infect conifers only (Larix decidua, L. kaempferi, and P. radiata). On the conifer hosts, spermatia and aeciospores are produced. The aeciospores reinfect the poplar hosts. Spread from conifer to conifer does not occur — the aeciospores produced on the conifer needles infect only poplars.
There are two patterns of dispersal related to the mode of winter survival. Where only telia survive, no rust appears on poplars until after aeciospores are dispersed from conifers. In the spring the teliospores produce basidiospores which infect the conifer host; aeciospores are subsequently produced on the conifer host. In warm areas where uredinia survive, new rust develops soon after leaves grow.
With the demise of the semi-evergreen Populus nigra 'Sempervirens' which allowed urediniospores to overwinter, inoculum build-up in spring has been considerably delayed. Most poplars grow a reasonable amount before the disease cycle of infection really gets going. Severity varies from year to year depending on the time of initial infection, presence of alternative hosts, and climatic conditions.
Mild wet weather favours infection of both conifers and poplars. At 18°C, more than 24 hours with free moisture on the needles are required for infection of Larix to occur.
NZ distribution: Melampsora larici-populina occurs throughout New Zealand; M. medusae died out but may occur in isolated pockets from time to time after reintroduction from Australia.
Economic impact: Initially devastating as Populus was an important shelterbelt and land stabilisation tree (Fig. 54). Since the arrival of the disease, alternative species have been used and a plant-breeding programme has developed resistant species.
Control: Rust-resistant poplar varieties have been planted and mixtures have been used to ensure genetic diversity as different races of the fungus have appeared. Measures include the application of chemicals, e.g., benodanil, triadimefon, bordeaux, copper oxychloride, and dodine, at 3-weekly intervals.
History: Melampsora larici-populina and M. medusae entered New Zealand in 1973 via trans-Tasman wind currents from Australia.
Melampsora larici-populina spread very quickly throughout the country but the incidence of M. medusa declined and 10 years later it could not be found at all. This decline is generally attributed to the lack of overwintering hosts. It is expected that there will be reintroductions from Australia from time to time.
References: Close et al. 1978; Forest Research Institute 1973; Spiers 1990; Wilkinson 1987, 2000.

Fig. 54: Populus hybrid shelterbelt defoliated by Melampsora larici-populina
This information is intended for general interest only. It is not intended to be a substitute for specific specialist advice on any matter and should not be relied on for that purpose. Scion will not be liable for any direct, indirect, incidental, special, consequential or exemplary damages, loss of profits, or any other intangible losses that result from using the information provided on this site.
(Scion is the trading name of the New Zealand Forest Research Institute Limited.)